")
")
What Do Orchids Eat?
Text and Photographs by Rick LOCKWOOD - from March 2013 in the AOS magazin "Orchids"
I began my orchid experiences in earnest in 2001. Most of my culture practices have been developed from the experiences and advice from more experienced growers. I tempered and refined these practices with my general knowledge of biology (especially ecology), which I had used to develop culture protocols for creatures like tropical fish and poison dart frogs.
In 1995, I began a career as an environmental toxicologist that focused my attentions on the internal mechanisms causing toxicity in aquatic organisms. I have been focused for the last six or so years on how the major ions (salts) cause imbalances and toxicity in both fresh and saltwater organisms and plants. The major ions influential in aquatic toxicity are the same critical ions for all plants, including orchids. The major ions are calcium (Ca), magnesium (Mg), potassium (K), sodium (Na), sulfate (SO4), bicarbonate (HCO3) and chloride (Cl). Together with nitrogen (N) and phosphate (PO4), these form the majority of what plant use as “nutrients.”
WHAT’S THE PROBLEM? I’ve focused primarily on slipper orchids, species of Paphiopedilum and Phragmipedium, but I also enjoy growing species of Phalaenopsis and Bulbophyllum. My first five or so years of orchid growing focused primarily on the physical aspects of orchid culture: getting down the light, temperature and humidity requirements of my plants. This is definitely the foundation of growing good orchids, and probably accounts for at least 75 percent of the success I’ve enjoyed at this point. But I’ve been plagued with regular chronic problems, and observed many common complaints and trends among different growers, that I now attribute to nutrition problems. Furthermore, in the case of orchids, nutrition cannot be completely separated from potting mix (Trelka, 2010) and water chemistry factors.
The laundry list of my problems and cultural contradictions include:
-
I could not grow Phalaenopsis in pots without killing them, although many growers have excellent results in either standard pot or semi hydro culture (where some roots may be continuously submerged in water without rotting). In general, I had much more success with epiphytic plants in baskets or on mounts than in any system of pots and potting mixes.
-
Plant leaves turning purple, even in low light.
-
“Easy” Paphiopedilum species like Paphiopedilum sukhakulii and Paphiopedilum callosum have been extremely difficult for me to keep alive for more than a few years, but blooming like crazy all the way to their demise.
-
Multifloral Paphiopedilum species only did well when only very small pots were used relative to the size of the plant. For instance, a 2-inch (5-cm) pot for a plant with a 24-inch (60-cm) leafspan. Increasing pot size generally precipitated root rot.
-
Root rot problems after short periods in bark or coconut husk chips (CHC) mixes. Short-term transfer to sphagnum would temporarily revive root growth, but extended periods in moss produced poor results.
-
I could use CHC mixes with multifloral Paphiopedilums (in small pots), but not long term with other Paphiopedilum species.
-
Use of calcium supplementing materials such as oyster shell or limestone did not offer long-term support of Paphiopedilums. But these materials are often a mainstay of other growers’ potting mixes for pH support.
-
Plants (mostly slipper orchids, but some other large and fast-growing orchids, such as a Phalaenopsis schilleriana and a Colmanera Wildcat), would grow quickly for a few seasons, followed by slow growth, stunted new growth, and susceptibility to disease until demise.
-
Seedlings out of flask initially grew very well up to a year, followed by lack of growth and susceptibility to disease until demise.
-
Some species where highly susceptible to Erwinia rots.
-
Leaf-tip burn in Phragmipedium.
Although these were the problems I was experiencing, I found that these were common for many fellow growers. Usually the excuse was, “Oh, species XYZ is known to be a tough one”, and then there are the contradictions of orchid cultural practices that conflict with the ecological circumstances of wild orchids:
-
“Rots occur due to watering at night.” But many orchid species get rained on at night or for days on end without rotting in the wild.
-
Paphiopedilum species in subgenus Brachypetalum are “sensitive to salts,” but five of the six primary Brachypetalum species are found so close to the ocean that they encounter salty ocean spray on a daily basis. Two of the most popular are Paphiopedilum niveum and P. leuchochilum
-
“Calcareous orchid species require calcium in the potting mix.” But many growers are successful with non-supplemented bark, CHC or inert mixes, and many calcareous species remain “difficult to grow” despite heavy limestone supplementation.
Using my research background I started intensifying my observation of other successful orchid grower’s practices, and reviewing research from the commercial, nonorchid, agricultural community to build a larger body of comparative data to look for patterns in success and failure.
One key source of information I discovered around 2004 came from Bob and Lynn Wellenstein’s AnTec Lab reading room, with the documents “Mineral Nutrition for Slipper Orchid Growers” (http://www.ladyslipper.com/minnut.htm ), and “Use of Coconut Husk Chips for Potting Medium” (http://www.ladyslipper.com/coco3.htm). Over the years, several articles in the reading room have been influential for my program. Two of the main points to take home from these two articles is the competitive, or antagonistic, interactions among potassium, calcium and magnesium, and the ion exchange capacity of coconut husk favoring the monovalent cations (sodium and potassium) in favor of the divalent cations (magnesium and calcium). At the time, however, I fairly discounted the notion of antagonistic or overdose interaction, because the predominant wisdom at the time was “orchids take what they need as you pour it on them, and leave what they don’t need.” This is the basic concept that agronomists used to develop feeding regimes based on leaf tissue mineral concentrations.
Please note, however, that the ratios of major nutrient ions listed in the WELLENSTEIN article on mineral nutrition for slippers are similar to the Michigan State Universty (MSU) fertilizer mix (or Arkenes Rain Mix), and the ratio of nitrogen to potassium is similar to just about all “balanced” commercial fertilizer mixes. Nutrient ratios based on leaf tissue concentrations of food crops and cultivated ornamental plants. Furthermore, literature was heavily focused on the N-P-K of a mix. Calcium and magnesium were only ascribed a limited or inconsequential role for non calcareous orchids.
With continued culture shortcomings after attempts to refine my fertilizing and potting practices based on my perceptions of what other successful growers were doing, I started experimenting with bone meal (calcium phosphate) and, later, Epsom salt (magnesium sulfate). These were supplementing practices that have been used for centuries in the general horticultural community, but had fallen out of favor in the orchid-growing community in favor of the more targeted MSU approach. Some short-term gains were quickly noted after calcium phosphate addition, primarily in returning some plants with purple leaves back to green. Much greater positive changes throughout my collection were noted after Epsom salt additions. Pale green or yellowing plants turned dark green, and leaf length increased by over 20 percent in some plants. The new growths and roots were larger and more durable. The patterning of the mottled-leaved paphiopedilums became distinctive, and in general the leaves of paphiopedilums became tougher and glossier. I was getting closer, but still having problems with roots, seedlings and occasional Erwinia bouts with large plants. So I dug deeper into the general agriculture and rainforest ecology literature (BRUIJNZEEL 1991, CROTHER 1987, MEDINA 1994, TURNER et al 2007, and VITOUSEK 1986).
THE DATA From these sources I deduced the following generalizations:
-
Plants in natural settings, regardless of species and habitat, have similar ratios of the primary elements. In priority Nitrogen > Calcium > Magnesium > Potassium > Phosphorus > Sodium. This includes an amazing array of plants from ocean kelp, river algae, and sphagnum moss to rainforest trees and shrubs.
-
Free, bioavailable potassium is relatively rare in the environment, forcing plants to actively uptake or recycle this nutrient when available.
-
Calcium and magnesium are relatively common in the soil and aquatic (liquid) environment so plant uptake of these nutrients is more passive (relative to potassium uptake).
-
Tropical orchids are primarily found in leaf litter, epiphytic on trees or lithophytic on cliffs or rocks. None of these habitats has large amounts of potassium relative to calcium and magnesium, and potassium is particularly rare in limestone lithophytic environments.
My specialty work in aquatic toxicology focusing on salt toxicity was also important to putting together the pieces of the puzzle. Because water flows downhill, knowing what is in the river, stream, creek or even ocean gives insight to what nutrition plants upstream in the trees or on cliff faces are accessing. The most obvious aspect of the aquatic toxicology of salt is that potassium is the most toxic element of the seven major ions, while calcium and sodium are the least toxic. Freshwater mussels, found worldwide, but globally imperiled in much of their present range, cannot handle much more than 10 to 20 parts per million (ppm) of potassium on a long-term basis, and 60 to 100 ppm will kill them in just a couple of days. The typical balanced fertilizer, applied at the rate of 100 to 200 ppm N will have just as much K, and is lethally toxic to this globally widespread aquatic organism. It is obvious that orchids in the wild do not experience weekly assaults of concentrated potassium. A typical stream, outside of agriculturally impacted areas where mussels are no longer found, will generally have 4 or less ppm of potassium. Rainwater rarely has any significant potassium, limestone is almost devoid of potassium and granitic feldspar is too insoluble to release significant amounts of potassium during a rain event. My house in Tennessee sits on a forested karst limestone ridge where native orchids can be found. A recent analysis of my well water showed 162 ppm Ca, 12 ppm Mg, 3 ppm sodium (Na) and less than 1 ppm K. Figure 1 shows the amounts of nitrate and K in throughfall, and drainage from epiphytic bird’s nest ferns in Sabah, Borneo. So where do orchids get their potassium?
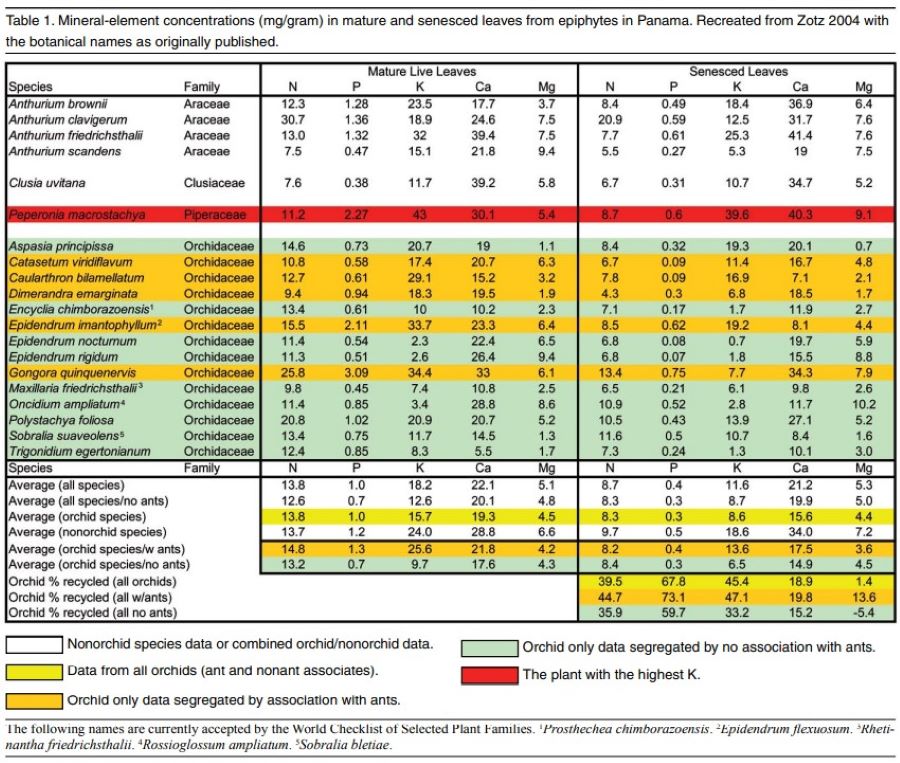
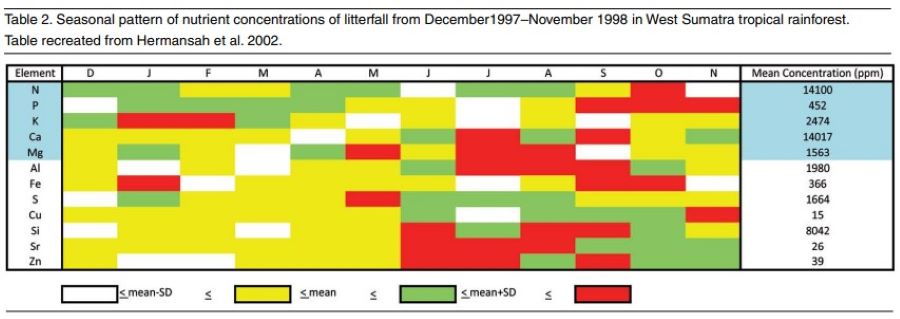
Looking at the tables from ZOTZ (2004) and HERNANSAH et al (2002) (Figures 2 and 3), we can see that the amount of K (and phosphorus too, for that matter) in plants is significant, and considerably more abundant than what is available from the air, water or the local geology. The situation for N, Ca, and Mg is different with respect to the amount of these materials in the local environment. However, in order for the K to reach the levels found in plant tissues, it must constantly recycle and bioaccumulate through the litterfall and soil making process. The Zotz work in particular demonstrates the importance of nutrient recycle of rare elements in the Panamanian rainforest, and shows that epiphytic plants (including orchids) reabsorb/recycle about half of their K during leaf senescence. Phosphorus (P) is recycled at a greater rate, but calcium and magnesium are recycled at low rates. A key factor I deduced from the ZOTZ work, but not mentioned in the text, is the relationship that ants have with some epiphytic plants and the nutritional role they play for these plants.
So noting that K is much rarer in the environment than in plant tissues, and also knowing that the MSU fertilizer is based on leaf tissue analysis, I started looking into plant physiology and cellular metabolism literature. Very little is written on wild orchids (or wild plants in general) with the bulk of literature based on studies with domesticated agricultural plants. I was able to summarize the following:
-
Epiphytic plants are very efficient at uptaking and sequestering K (Winkler 2010). Apparently this is also an energy requiring process. The uptake of K is like the “kids in the candy store” analogy. Wild epiphytic plants do not have the capacity to regulate the uptaking and storing of K in their tissues.
-
Given the active transport of potassium in plants, bioavailable potassium will be readily taken up by plants in excess, which will then ultimately block the uptake of calcium and magnesium (antagonism, Figure 4, Poole and Seely 1978). This will in turn reduce the uptake of phosphate (which is facilitated by the presence of calcium and magnesium in plant tissues, Shaibur 2008).
-
Plants that have incurred unfavorable tissue ratios of potassium/calcium/magnesium will have reduced growth and are susceptible to disease (Easterwood 2002).
-
The regular use of high-potassium fertilizer is indicated for fast-growing plants that are completely harvested at the end of a single growing season (like corn, wheat or rice). Slow growing perennial crops (such as fruit trees) need only intermittent potassium supplementation during the fastest annual vegetative growth phase, and when soil is depleted (Thompson).
THE SOLUTION Armed with the above information, I needed a new fertilizing strategy for all of my orchids, and a new potting strategy for the paphiopedilums. I first compared the proportional ratios of the major ion content (NPK, Ca and Mg) of what I was using with what the wild plants contained in either live or senesced leaves.
-
GreenCare MSU (pure water, %) 13-3-15-8-2 OrchidmixTM MSU 12-6-13-7-2
-
Zotz, 2004 (average all species, live leaves, mg/gr) 14-1-18-22-5
-
Zotz , 2004(average Encyc, Epis and Onc only, senesced leaves, mg/gr) 8–0.2–2–15–7
-
Zotz, 2004 (average non ant orchids, senesced leaves, mg/gr) 8 – 0.3 – 6.5 – 15 – 4.5
-
Hermansah, 2002 (leaf litter, mg/gr) 14-0.4-2.4-14-2
-
Naik, 2007 (live leaves, mg/gr) 4 – 1 – 9 – 24 – 13
Other leaf litter data are within the ranges of the ratios listed above, but some trends were obvious. Excluding the data from ant-fed plants, the amount of potassium is generally less than half the calcium and no more than equal the amount of magnesium. In leaf litter or senesced leaves the amount of potassium is less than the amount of nitrogen. Because the plants are recycling a certain amount of nutrients (mostly the N-P-K), I considered the leaf litter and senesced leaf data to represent what the plants were actually loosing, and what they needed for replacement for either maintenance or growth. Furthermore, the orchids living in leaf litter accumulations on the forest floor (such as some of my favorite Paphiopedilum species) would be experiencing the nutrient concentrations found in the mixed leaf litter as their primary nutrient source. Thirdly, the paper on the uptake of potassium by epiphytes (Winkler 2010) indicated that epiphytes would obtain sufficient potassium from very dilute sources, which I interpreted as natural surface water concentrations not toxic to aquatic life.


In a similar fashion to my work with freshwater mussels, I developed a new fertilizer mix by substituting available salts with different cationic makeup. My reduced K, increased Ca/Mg program was initially accomplished by reducing the amount of MSU fertilizer by half, and adding equal parts of calcium nitrate and magnesium sulfate. This kept the nitrogen levels constant while drastically reducing K and increasing both the calcium and magnesium. Ultimately, I cut this weekly total amount of feeding in half again so total nitrogen input was 30-50 ppm, only occasionally going back up to 100 ppm on warm sunny days. The expected nutrient ratios for this fertilizer mix were: 6-3-3-8-4 if applied at roughly 50 ppm N. Between weekly fertilization events, all irrigation water is supplemented with calcium and magnesium (primarily through the addition of my well water diluted 10:1 with reverse-osmosis [RO] water). As I noted in my research, growers utilizing surface or tap waters to mix up their fertilizer (as opposed to RO-, distilled- or rainwater), were less apt to experience the symptoms I discussed earlier. Also note that if MSU pure water formula is mixed in a medium hard or hard surface water, then the resulting K:Ca:Mg ratios are similar to the Zotz 2004 all species live leaf data. Epsom salt was supplemented intermittently in regular irrigation water on warm, sunny days.
I began this program in spring 2011, but by winter 2011, a collaboration of my fellow Slippertalk members, spearheaded by Ray Barkalow (First Rays), Doylestown, Pennsylvania, working with GreenCare, produced another formula we call “K-Lite.” For a use rate of roughly 100 ppm N the nutrient ratio is 12-1-1-10-3 if added to RO water. Because I mix it at roughly 50 ppm N in soft water (which has additional Ca and Mg), my final mix for 2012 and going into 2013 would be roughly equivalent to a 6-0.5-0.5-6-2 ratio. Using the mixture at 50 ppm N, I would still be offering 4 ppm of K, which appeared more than sufficient for my orchids. The K-Lite was made by GreenCare, who also mix the standard MSU formula. The K-Lite formula contains all the same trace metals as the standard MSU formula. This basic inorganic feeding regime was (and still is) occasionally supplemented with a liquid kelp extract (Seaplex from Botanicare) at the rate of ¼ tsp per gallon. Kelp products also contain macronutrients NPK Ca and Mg, micronutrients, a host of amino acids, and small amounts of plant hormones. At ¼ tsp per gallon, the amount of K added is less than 3 ppm. As noted, my use of kelp is intermittent, based on my impression of rate of growth, and other growing conditions. More evolution of my feeding regime was yet to come as new in situ research data was obtained, and I was able to correlate new and changing growth observations with potting systems.

As I noted in the Problems section, my choice of potting system was influential to my success with my epiphytic species, so I attempted to apply that experience to my slippers. My early attempts at using a more epiphytic potting system (baskets) for slipper culture showed promise for a few months. However, after roughly six months, some plants started loosing ground again, and I began to put “two plus two” together with regard to coupling K toxicity with the variables of potting matrix effects on the concentration exposure of the fertilizer components. My basic observation and theory was: per given concentration and application frequency of fertilizer, the mounted plants would experience the lowest total potassium over time, while the potted plants in the most retentive / high Cation Ion-exchange Capacity (CEC) materials would experience the greatest “dose” of K over the same time frame. Figure 5 (data from Xavier Garreau) shows how influential potting mix selection can be on the uptake rate of K in orchids in support of my theory. By Spring/Summer of 2013, I was putting all this together, and encouraged by the research of Turner et al (2007), I further reduced the concentration of fertilizer to approximately 5 ppm N. However, I increased the application frequency to every watering, which was daily for mounted plants, every 2-3 days for plants in baskets, and every 3-5 days for potted plants. Also noting a handful of plants in higher light levels were getting pale leaves, I added a supplement of magnesium/phosphate/sulfate I concocted. So the final concentrations of my nutrient regime, now synonymous with my basic irrigation water, (and with no kelp additive) were:
-
Nitrate N = 4.5 ppm
-
Ammonia N = 0.17ppm
-
Phosphate (PO4) = 2.2ppm (or 1.7 ppm as P2O5)
-
Potassium <1ppm
-
Hardness = 20ppm as CaCO3 (calcium ~ 4.5ppm, and magnesium~2.5ppm)
-
Sulfate = 3ppm
-
Alkalinity = 12ppm as CaCO3
-
pH = 6.87 s.u.
-
Conductivity = 49 µS/cm
So even with the more open basket system, I still needed to reduce both the relative amounts of high CEC components and/or reduce the overall concentration of applied nutrients. The present setup for my slippers in baskets is to line them with sphagnum moss and fill up most of the space with large (approximately 1” diameter) limestone gravel (this is too large and dense to significantly alter chemistry). Additional moss, bark, or coconut husk mix may be added to fill in some more gaps and retain a bit more moisture, but I try to keep these materials down to a minimum, or be prepared to replace them after a few months if roots do not make it to the outside of the baskets. I do not use calcium carbonate supplements any more. Feeding at the very dilute rates I feed at now does not significantly reduce pH in the baskets. Although I am very pleased with the results of the basket system, I have also been seeing a resurgence of good results in traditionally potted bark or CHC based mixes in various types of pots. This reinforces my belief that pot chemistry is optimized at very low application rates of inorganic nutrients.
It is now the beginning of 2014 with two and a half years of this low K/high Ca-Mg fertilizing regime I am pleased with the results. Leaves are larger, stiffer and shinier. Erwinia, mealybug and scale problems are greatly reduced. Plants that were in decline are rebounding. Seedlings are transitioning out of compots into individual pots without stalling and dying. Five of my plants even received AOS quality awards in the last year (two were raised from my own breeding). I realize that two and a half years is a small time frame with regards to orchid culture so time will tell if these short term results are meaningful. In the meantime, I’ll be assessing and tweaking. The Zotz 2004 data showed the influence of ants on the nutritional status of their tended plants. I only have a small number of ant-favored species in my collection (such as Gongoras and Coryanthes), so they may warrant closer attention and maybe some additional K during their fast growth periods. Watching, testing, trying new things.
What a glorious hobby!
Addendum:
in September 2022 i was in contact with Rick LOCKWOOD again:
He has confirmed to me that he is still using the K-Lite fertilizer (which celebrated its 10th anniversary in December 2021) as a base feed at a rate of 5-8ppm N per day as a daily watering/fertilizing. I also add a squeeze of lemon juice and a small amount of seaweed extract and potassium silicate.
K-Lite fertilizer is difficult to get in the EU, but you can easily make it yourself (see https://firstrays.com/do-it-yourself-k-lite/).
To do this, take 1/3 weight of the Rain Mix fertilizer from Akerne + 1/3 Calcinit (Calciumnitrate) + 1/3 Epsom salt. This fertilizer formula is then: 9.1 - 0.9 - 4.6 - 10.3 - 4.5